神谷研では、生命の基本単位である細胞膜に着目し、人工細胞を構成する膜構造、膜タンパク質機能、細胞らしいシステムの再構成に取り組んでいます。細胞膜は、脂質非対称性、膜サイズ、階層構造、膜張力などの物理化学的特徴をもつ動的な界面であり、生命機能を生み出す重要な場です。私たちは、まず人工膜を精密につくり、その膜にナノポアやチャネルなどの膜タンパク質を組み込むことで、物質輸送や分子認識の仕組みを明らかにしています。さらに、無細胞タンパク質合成、酵素反応、刺激応答、成長・分裂を統合し、生命らしい振る舞いを示す人工細胞システムの構築を目指しています。これらの研究を通じて、生命現象の理解と、創薬・医療・バイオものづくりへの展開を進めています。
細胞膜の基本構造を再構成する
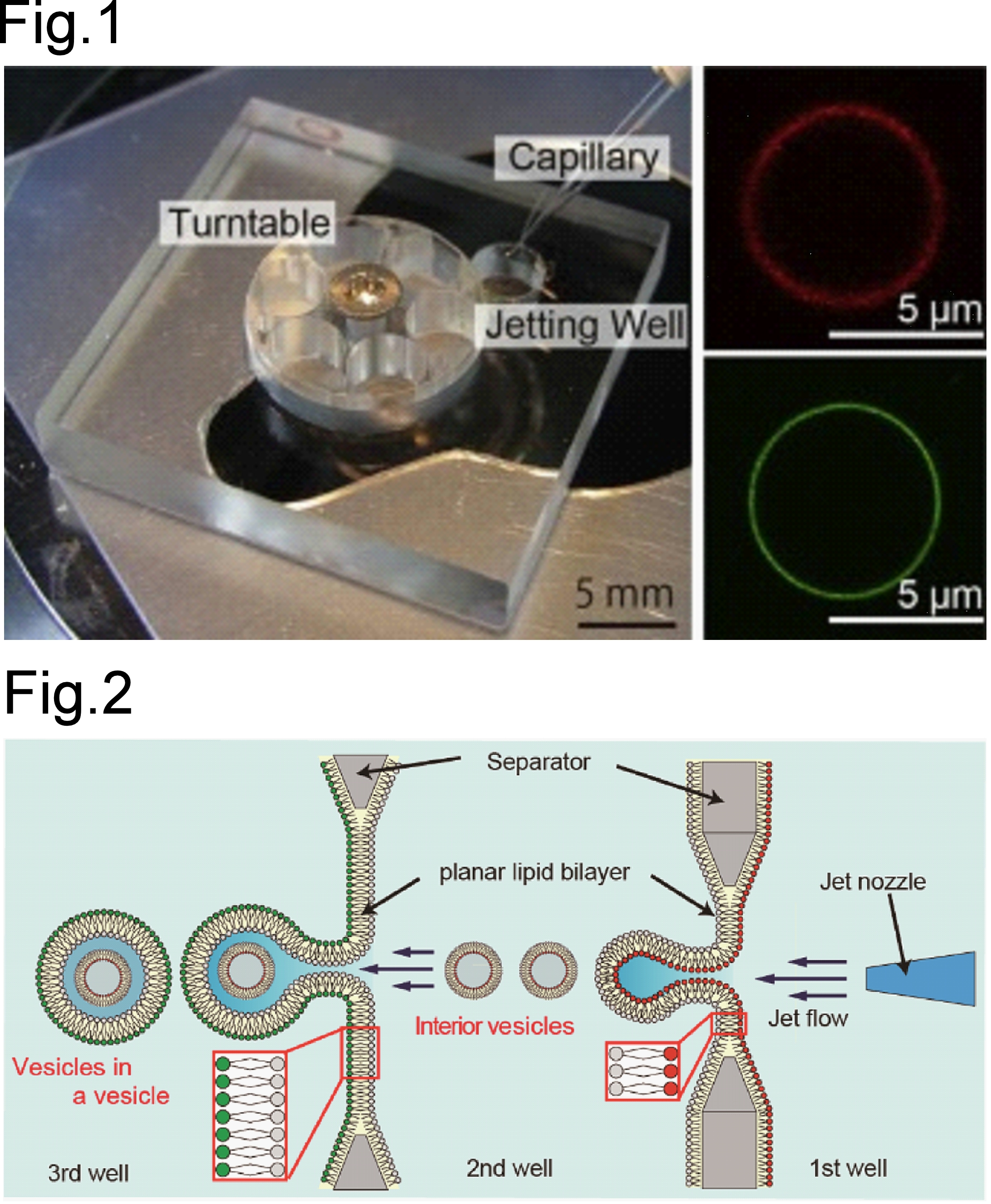
細胞膜は、単なる脂質二重膜ではなく、内外で脂質組成が異なる非対称性、細胞サイズの閉じた構造、小胞を内包する階層性、膜張力や曲率といった物理化学的特徴をもつ動的な分子集合体です。神谷研では、細胞膜の基本構造を人工的に再現するため、MEMS技術とパルスジェット法を用いて、有機溶媒の影響を抑えた細胞サイズ非対称脂質ベシクルを作製し、非対称膜を人工膜上で解析する基盤を築いてきました (Fig.1) [1]。さらに、非対称脂質ベシクルを連続的に生成する技術を発展させ[2]、細胞サイズ非対称脂質ベシクルの内部に小型の非対称ベシクルを含むvesicles-in-a-vesicleを構築することで、細胞膜と細胞内小胞膜の階層構造を模倣しました (Fig.2) [3]。また、パルスジェット法をナノサイズ領域へ展開し、50–300 nm程度の非対称脂質ナノベシクル形成にも成功しています[4]。さらに、完全な人工膜である外膜:リン脂質、内膜:両親媒性タンパク質から形成される非対称膜ベシクルの構築に成功しています[5]。これらの研究により、人工細胞を支える「膜の器」の形成原理、サイズ制御、膜物性、階層構造を明らかにし、膜タンパク質や反応系を組み込むための基盤技術を構築しています。
[1] Kamiya et al., Nat. Chem. Vol.8 (2016) pp.881-889.link
[2] Gotanda et al., Sens. Actuators B Vol.261 (2018) pp.392-397.link
[3] Kamiya et al., RSC Adv. Vol.9 (2019) pp.30071-75.link
[4] Kamiya et al., Sens. Actuators B Vol.327 (2021) 128917.link
[5] Suzuki and Kamiya, iScience Vol.26 (2023) 106086.link
神谷ら、生物物理 (2018) Vol.58 No.6 pp.291-296.
神谷ら、生化学 (2018) Vol.90, pp.225-229.link
膜タンパク質を人工細胞膜で機能させる
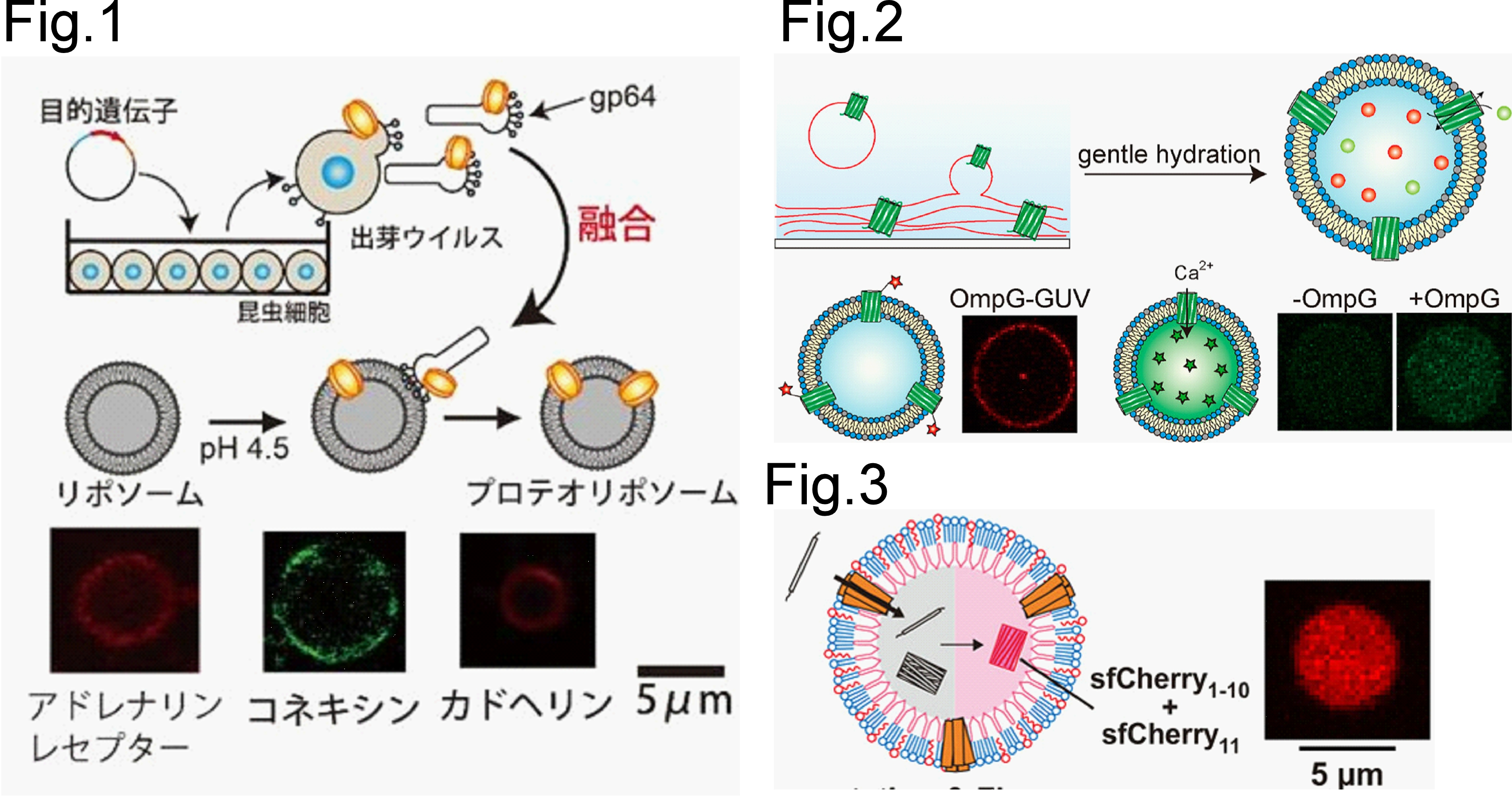
膜タンパク質は、細胞膜を介した物質輸送、分子認識、環境応答、情報伝達を担う生命機能の中心的な分子です。しかし、膜タンパク質は脂質膜環境に強く依存して折りたたまれ、機能するため、人工細胞膜上でどのように合成し、組み込み、働かせるかが重要な課題です。バキュロウイルスの膜融合能を利用したプロテオリポソーム作製法を確立し、アドレナリンレセプター[1]、コネキシン[2]、カドヘリン[3] 等の膜タンパク質の機能観察に成功しています (Fig.1) 。特に、カドヘリンリポソームは、ガン細胞に特異的に結合し、カドヘリンリポソームは効率的に細胞内へ取り込まれることを発見しました。OmpGやOmpAなどのグラム陰性菌外膜タンパク質を無細胞翻訳系で合成し、脂質膜へ挿入・再構成する条件を解析してきました。特に、大腸菌膜に近い脂質組成をもつリポソームでOmpGの膜挿入が促進され、パッチクランプ解析により機能的なナノポア形成を確認しました[4]。また、乾燥脂質膜–膜タンパク質フィルムの再水和により、改変OmpGを細胞サイズリポソームへ組み込む手法を開発し、人工細胞膜上で膜タンパク質を機能させる技術を発展させています (Fig.2) [5]。さらに、機械刺激応答チャネルMscLを非対称脂質–両親媒性タンパク質ベシクルに再構成し、膜張力の変化を分子輸送へ変換できることを示しました (Fig.3) [6]。これらの研究を通じて、人工細胞膜上で膜タンパク質を作り、働かせ、測ることで、生命膜の機能原理を理解し、人工細胞への輸送機能付与へ展開しています。
[1] Kamiya et al., Biochim. Biophys. Acta – Biomembranes Vol.1798 (2010) pp.1625-1631.link
[2] Kamiya et al., Biotechnol. Bioeng., Vol.107 (2010) pp.836-843. link
[3] Kamiya et al., Biomaterials, Vol.32 (2011) pp.9899-9907. link
[4] Kamiya Sci. Rep., Vol.12 (2022) 2376. link
[5] Tosaka and Kamiya, ACS Omega, Vol.9 (2024) pp.5911-18. link
[6] Baba and Kamiya, ACS Appl. Mater. Interfaces, Vol.16 (2024) pp.21623-32. link
タンパク質ナノポアを設計し、生体分子を検出する
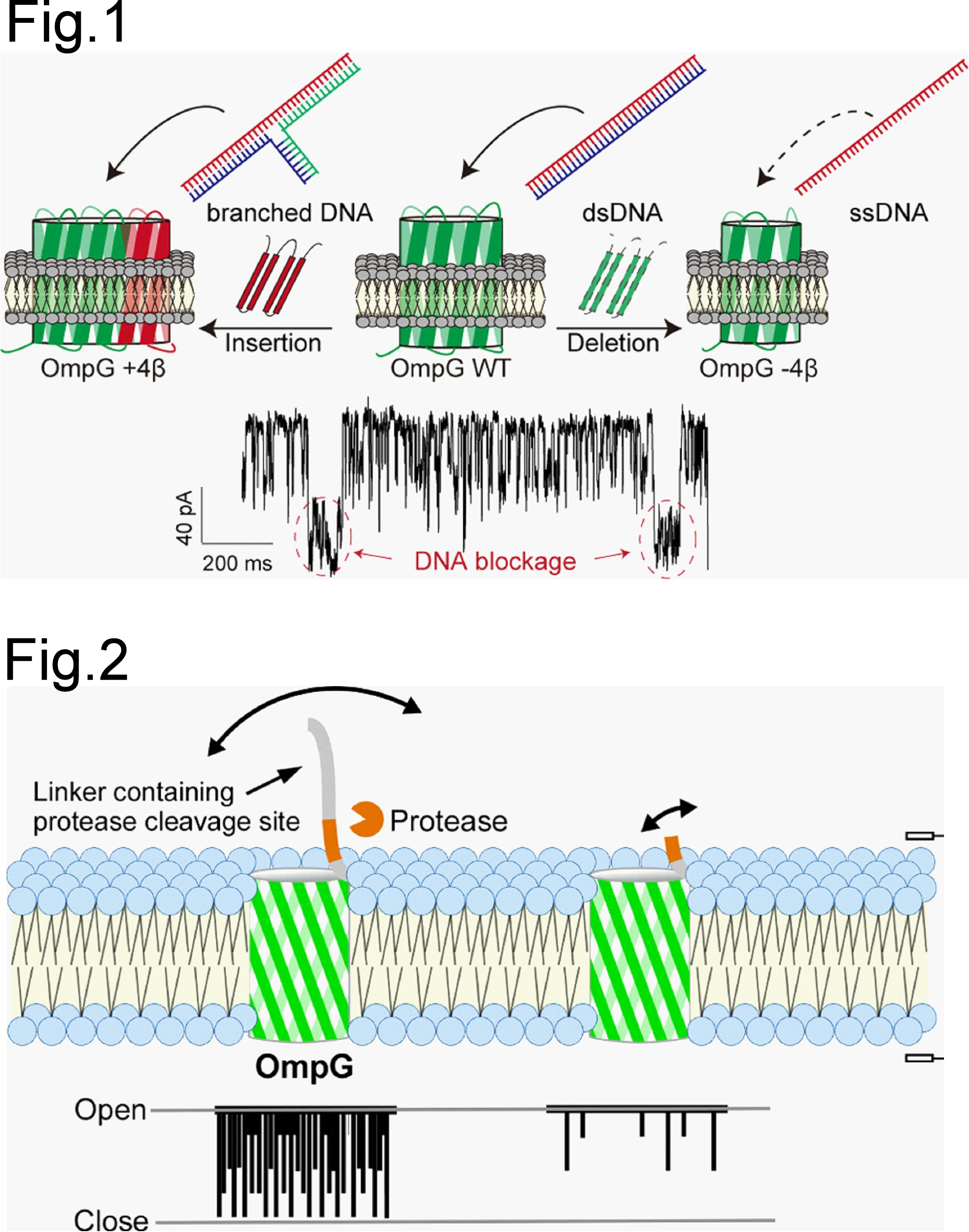
タンパク質ナノポアは、脂質膜を貫通するナノメートルサイズの孔を通じて、イオンや分子の通過を電流変化として1分子レベルで検出できる分子センサーです[1]。神谷研では、βバレル型膜タンパク質OmpGを中心に、ナノポアの構造改変と機能設計に取り組んでいます。OmpGのβヘアピン構造を拡張・短縮することで、野生型とは異なる孔径やゲーティング特性をもつナノポアを作製し、二本鎖DNAとの相互作用をイオン電流から識別できる系を構築しました (Fig.1) [2]。また、OmpGのN末端に導入するリンカー長を変えることで、ポアの開閉挙動を制御し、TEV protease、thrombin、HRV3C proteaseなどによるリンカー切断を電流変化として検出する系へ展開しました (Fig.2) [3]。さらに、磁気ビーズとリポソーム膜上のナノポアを組み合わせ、精製を必要としないmicroRNA検出系も示しています[4]。現在は、βバレル型膜タンパク質を構造設計の出発点とし、より大きな孔径、安定な電流特性、分子認識能をもつ新しいタンパク質ナノポアの開発にも挑戦しています。
[1] Kamiya et al., Sci. Rep., Vol.8 (2018) 17498. link
[2] Tosaka and Kamiya, ACS Appl. Nano Mater. Vol.5 (2022) pp.6149-58.link
[3] Kamiya et al., ACS Appl. Bio Mater., Vol.9 (2026) pp.3613-21. link
[4] Fujii et al., Anal. Chem., Vol.90 (2018) pp.10217-22. link
外部刺激に応答する人工細胞システムを設計する
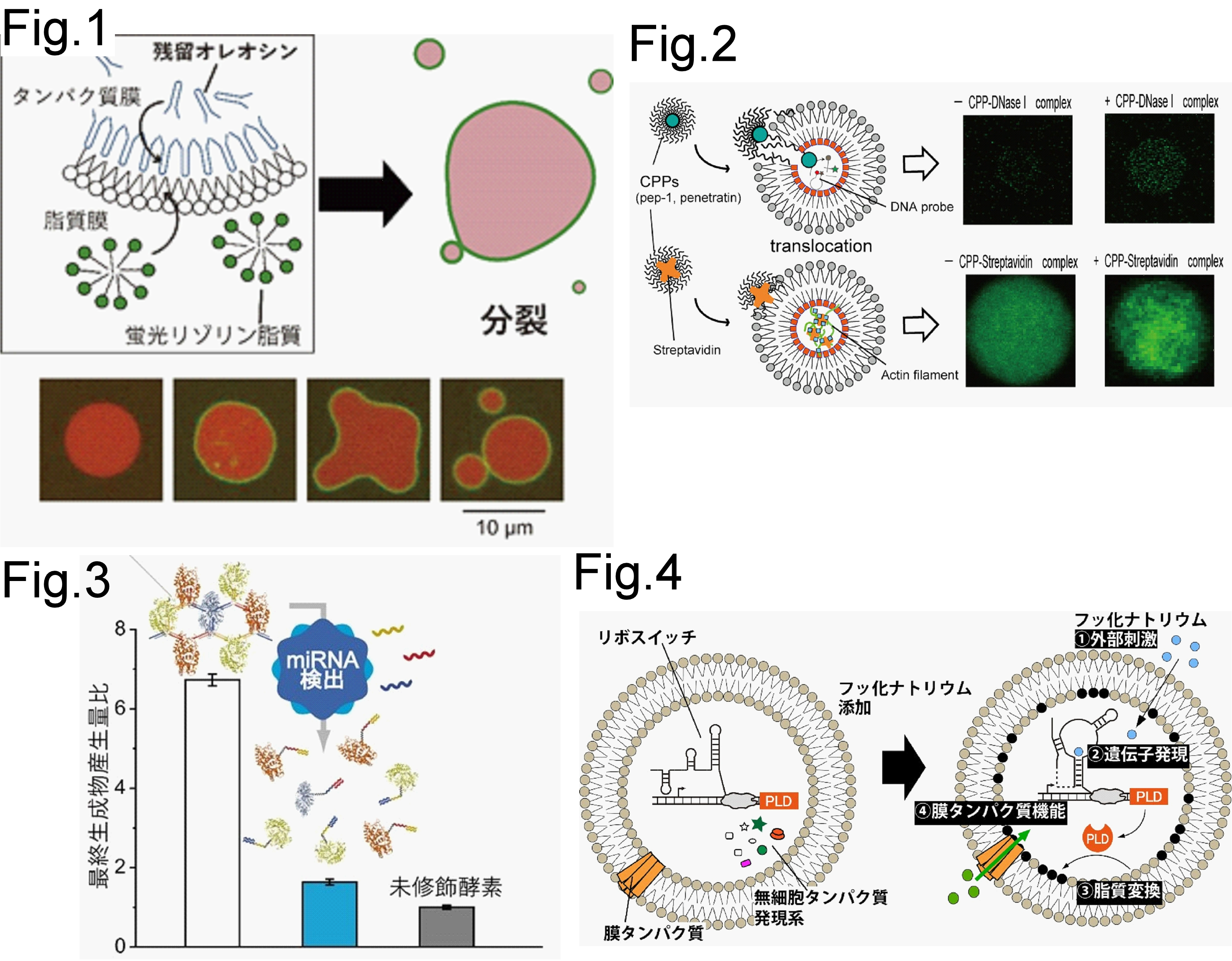
人工細胞は、脂質膜、膜タンパク質、遺伝子発現、酵素反応、分子輸送を統合し、細胞らしい振る舞いを人工的に再構成する研究です。神谷研では、膜を単なる容器として扱うのではなく、外部刺激を受け取り、内部反応や膜機能を変化させる動的なシステムとして設計しています。外膜ホスホリパーゼを利用した二機能性リポソームでは、小分子輸送と小胞出芽を組み合わせた人工細胞モデルを構築しました[1]。また、オレオシンを用いたリン脂質–両親媒性タンパク質非対称膜ベシクルでは、膜タンパク質の再構成、ベシクルの成長、リゾ脂質添加による分裂、分子輸送を示す人工細胞モデルを実現しました (Fig.1) [2]。非対称膜ベシクルの内膜を形成するオレオシンに光や薬剤で水溶性タンパク質の集積に成功しています[3]。さらに、細胞透過性ペプチドを用いてリン脂質非対称膜リポソーム内部へタンパク質を輸送し、内部の酵素反応やアクチンネットワーク形成を任意のタイミングで開始させる系を開発しています (Fig.2) [4]。加えて、脂質組成が細胞サイズベシクル内の無細胞タンパク質合成時の転写・翻訳に与える影響を解析し[5]、DNA computingを統合した酵素ネットワーク制御にも展開しました (Fig.3) [6]。最近では、リボスイッチ制御によるホスホリパーゼD発現と脂質変換を組み合わせ、外部刺激に応答して脂質変換と膜タンパク質機能を誘発させる人工細胞システムを構築しています (Fig.4) [7]。
[1] Ohnishi and Kamiya, ACS Synth. Biol. Vol.10 (2021) pp.1837-1646.link
[2] Suzuki and Kamiya, iScience Vol.26 (2023) 106086. link
[3] Suzuki and Kamiya, ChemPlusChem Vol.91 (2026) e70164. link
[4] Miwa and Kamiya ACS Synth. Biol., Vol.11 (2022) pp.3836-46. link
[5] Miwa et al., Sci. Rep., Vol.14 (2024) 2852. link
[6] Mameuda et al., Anal. Chem., Vol.95 (2023) pp.9548-54. link
[7] Kamiya et al., Commun. Biol., Vol.9 (2026) 580. link
Suzuki and Kamiya, ChemSystemsChem, Vol.8 (2026) e00071. link
神谷、豊田, ファルマシア, Vol.64 (2025) pp.319-23.
化学・生物学分野へ応用するマイクロデバイスの開発
微細加工技術を用いることによりマイクロサイズの微小流路を作成することが可能です。微小流路は、体積よりも表面積の効果が支配的になるので、反応や温度制御を迅速に正確に行うことができます。近年、化学や生物分野の研究で広く用いられている技術です。
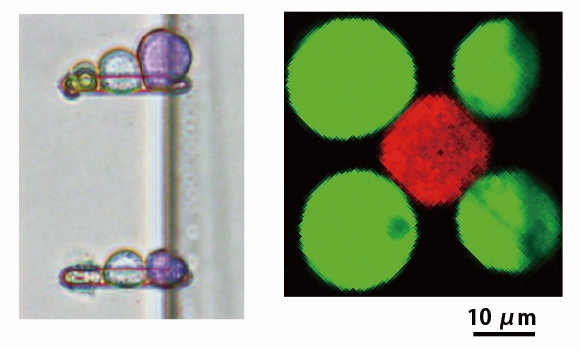
対数濃度勾配を作成可能なマイクロデバイスを考案し、膜タンパク質の薬物阻害曲線を簡便に作成することが可能になりました[1]。静水圧を利用して、細胞を任意の順番や形状で並べるデバイスを考案しています(図)[2]。このデバイスは、空間的に細胞を制御した状態で細胞間相互作用が観察でき、詳細な細胞間相互作用の理解が可能になります。
リン脂質組成非対称膜リポソームの作成においてもマイクロデバイスを利用しています。
[1] Abe and Kamiya et al., Analyst, Vol. 140 (2015) pp.5557-5562.link
[2] Kamiya et al., Advanced Healthcare Materials, Vol.7(2018)1701208. link